研究内容
私達の体の構成単位である細胞はさらに様々な膜系いわゆる細胞内小器官(オルガネラ)により構成されていますが、これらのオルガネラは決して独立した器官ではなく、小胞(膜)輸送を介して情報交換を行っています。適切な膜交通は生命活動の維持に不可欠なことから、その制御メカニズムの解明は生物学・医科学における重要な課題の1つと考えられます。適切な膜交通を行うためには、交通整理人の存在が重要で、これまで数多くの制御分子の存在が明らかになっています。私達の研究室ではシナプトタグミンと低分子量GTP結合蛋白質Rabという二つの交通整理(膜輸送制御)因子に焦点を当て、これらの分子ファミリーによる膜輸送制御の分子メカニズムの解明に取り組んでいます。これまでの研究で、調節性分泌(例えば、シナプス小胞からの神経伝達物質の放出)におけるシンプトタグミンやRab27の役割とメラニン色素輸送の分子メカニズムの解明に成功しています。
(1)調節性分泌におけるシナプトタグミンフミリーの役割
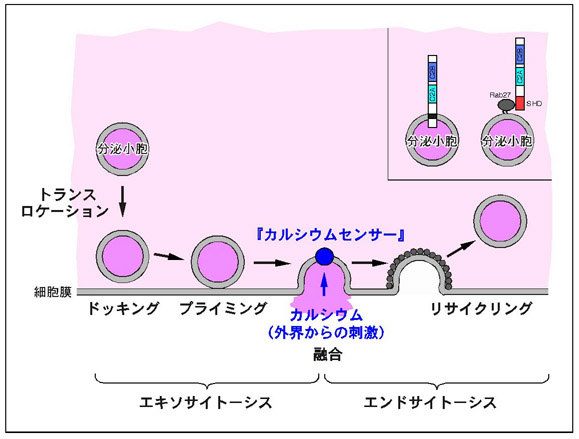
細胞は外界からの刺激に対応して様々な物質を放出する(この過程を「分泌」と呼びます)ことにより、個体としての生命活動を維持しています。分泌には細胞が常時行っている構成性の分泌と外界の刺激に依存する調節性の分泌に分けることが出来ます。例えば、神経細胞からの神経伝達物質の放出や内分泌細胞からのホルモン分泌がこの範疇に入ります。「分泌」と言う僅か二文字で書けてしまう生命現象ですが、実際には図1に示したように少なくとも四つのステップからなる複雑な過程により構成されています。シナプス小胞の輸送を例にとると、シナプス小胞はまずプレシナプスの先端部まで運ばれプレシナプス膜に結合します(ドッキングのステップ)。ATP依存的なプライミングという準備段階を経て、シナプス小胞はプレシナプス膜と融合し(フュージョンのステップ)、内容物である神経伝達物質が細胞外へと放出されます(これらの過程をまとめて、開口放出あるいはエキソサイトーシスと呼びます)。放出された神経伝達物質や制御因子は再利用され、次の神経伝達物質の放出に備えます(リサイクリングのステップあるいはエンドサイトーシスと呼びます)。シナプス小胞などの分泌小胞と細胞膜の融合は細胞内のカルシウムイオン濃度により厳密に制御されていることから、分泌小胞上にはカルシウムイオン濃度の上昇を感知する「カルシウムセンサー」の存在が提唱されています。
(詳細は本文を参照)
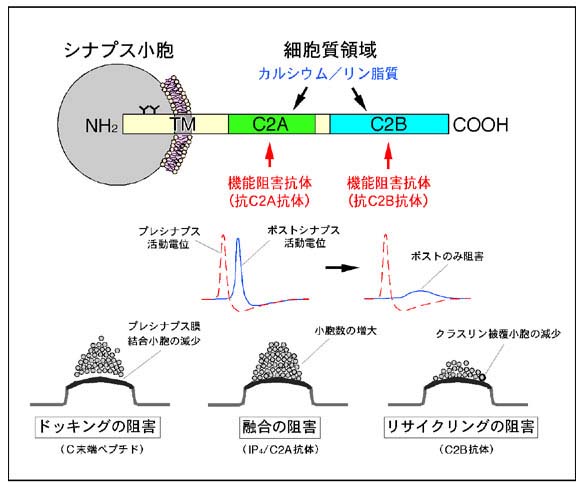
シナプス小胞の場合には、カルシウム・リン脂質結合蛋白質として知られているシナプトタグミンI分子が神経伝達物質放出の際のカルシウムセンサーの最有力候補と考えられています。シナプトタグミンIは図2に示すように、N末端側に膜貫通領域を一箇所、C末端側の細胞質領域にはプロテインキナーゼC(PKC)のC2調節領域と相同性を有する配列が二箇所タンデムに存在しています(それぞれC2A、C2Bドメインと呼ばれています)。私達の研究室では、1994年にイノシトール1,3,4,5-四リン酸(IP4)受容体としてシナプトタグミンII/IP4BPを同定したのをきっかけに(J. Biol. Chem. (1994) 269, 29206-29211)、シナプトタグミンI及びIIの神経伝達物質放出における機能解明に取り組んでいます。これまでのイカの巨大軸索を用いた解析から、シナプトタグミンI分子は単なるカルシウムセンサーではなく、シナプス小胞のドッキングの過程(PNAS (2000) 97, 14715-14719)やリサイクリングの過程(PNAS (1995) 92, 10708-10712; PNAS (2004) 101, 17855-17860)を制御する多機能分子であることを明らかにしています(図2)。特に、C2AドメインとC2Bドメインはアミノ酸レベルで40%以上の相同性を示すにも関わらず生化学的には全く異なる機能を有すること(J. Biol. Chem. (1995) 270, 26523-26527)、二つのドメインはお互いに協調し合いながらもシナプス小胞輸送過程において異なる機能を持つこと、C2Bドメイン内には複数のリガンド結合サイト(エフェクタードメイン)が存在することを明らかにした点は、当該研究分野において高く評価されています。
図2 シナプス小胞輸送におけるシナプトタグミンIの機能図 (クリック)
シナプトタグミンIは神経伝達物質放出の際のカルシウムセンサーとして機能する
以外にも、シナプス小胞のドッキングやリサイクリングのステップにも関与する多機
能分子と考えられています。例えば、C2Bドメインに対する抗体はシナプス小胞のリ
サイクリングのステップを顕著に阻害しますし、シナプトタグミンIのC末端ペプチ
ドはシナプス小胞のドッキングのステップを阻害します。
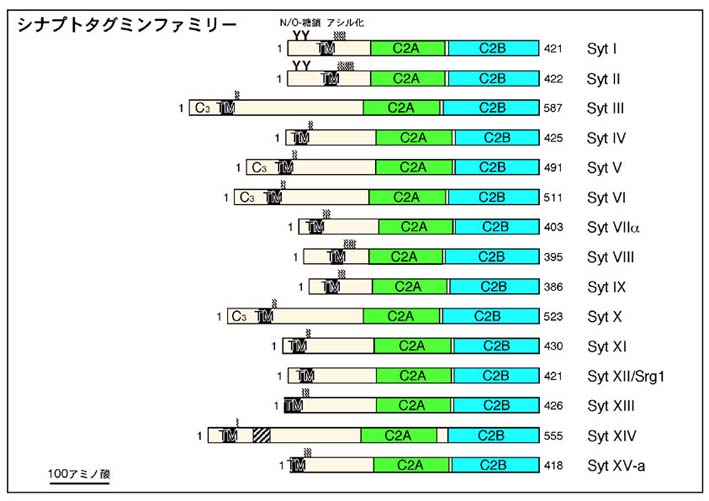
シナプトタグミンファミリーは現在では、植物、無脊椎動物(線虫、ショウジョウバエなど)、脊椎動物を含む幅広い生物種に存在し、進化的にも保存された蛋白質ファミリーです。マウスやヒトにおいては、これまで15種類の異なるアイソフォームの存在が報告されており(図3)、シナプトタグミンI同様、何らかの調節性分泌の制御に関わるものと考えられています。これまで、PC12細胞や膵臓β細胞などの内分泌細胞からのホルモン分泌においてはシナプトタグミンIXが(J. Biol. Chem. (2002) 277, 4601-4604)、グリア細胞の一種であるアストロサイトからのグルタミン酸放出にはシナプトタグミンIVが(PNAS (2004) 101, 9441-9446)、授精時におこる精子先体反応においてはシナプトタグミンVIが(Dev. Biol. (2001) 235, 521-529)重要な役割を果たしています。これに対し、10番以降のシナプトタグミンアイソフォームの機能は現段階では全く解明されおらず、これらのアイソフォームがどのような調節性分泌の過程に関与しているのかを今後明らかにして行きたいと考えています。
図3 マウス・シナプトタグミンファミリーの構造 (クリック)
シナプトタグミンは進化的にも保存された一つの大きなファミリーを形成し、マウ
スやヒトにおいては15種類の異なるアイソフォームの存在が報告されています。シナ
プトタグミンアイソフォームはいずれもN末端側に膜貫通領域を一箇所、C末端側に
タンデムC2ドメインを有しています。
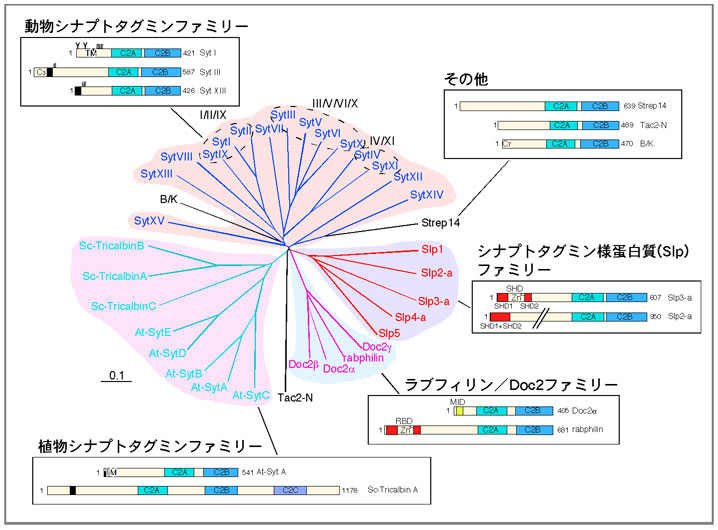
シナプトタグミンに代表されるタンデムC2蛋白質群は、植物、無脊椎動物、脊椎動
物に至るまで幅広く保存されており、マウスにおいては27種類もの異なる分子が存在
することが知られています。これらの分子は、アミノ末端側の構造により、幾つかのフ
ァミリーに分類されています。シナプトタグミンファミリーは膜貫通領域を持ち小胞上
に直接埋め込まれており、シナプトタグミン様蛋白質(Slp)は低分子量G蛋白質Rab27
を介して小胞上に存在します。
(2)調節性分泌におけるSlp(シナプトタグミン様蛋白質)ファミリーの役割
調節性の分泌は、何も神経系に限った現象ではなく、体中の様々な細胞で見られる現象です。上述のシナプトタグミンは主に神経系で強い発現が見られることから、非神経系においてはシナプトタグミンに類似した分子が機能している可能性が考えられます。私達の研究室では、2001年にマウス及びヒトのゲノムに存在するシナプトタグミン類似分子の探索を網羅的に行い(具体的には、シナプトタグミンと同様にタンデムC2ドメインを持つものをスクリーニング)、新規蛋白質ファミリーを含む27種類のカルボキシル末端型(C-タイプ)タンデムC2蛋白質を同定することに成功しました(図4)。この過程で、シナプトタグミンファミリーとDoc2/ラブフィリンファミリーとは系統樹上明らかに異なる枝(ブランチ)を形成する新規の蛋白質として、シンプトタグミン様蛋白質(synaptotagmin-like protein; Slp(スリップ)と略)を同定・命名しました(BBRC (2001) 281,
1226-1233)。Slpファミリーはマウス及びヒトにおいては5種類のアイソフォーム(図5)(Slp1/JFC1、Slp2-a、Slp3-a、Slp4/granuphilin、Slp5)が、ショウジョウバエにおいては1種類のアイソフォーム(dm-Slp/Btsz)の存在が報告されています。Slpファミリーの構造的な特徴は、シナプトタグミンファミリーと異なりN末端側に膜貫通領域の代わりに、Slpファミリーでのみ保存されたSlp homology domain(SHD領域)を有する点です(BBRC (2001) 283, 513-519)。SHD領域はタンデムC2ドメインを持たないSlac2(Slp homologue
lacking C2 domains)と命名した新規の蛋白質ファミリーにも見いだされ(図5)、SHD領域は何らかの蛋白質モチーフではないかと考えられました。SHD領域を見いだした当初は、この領域の機能は全く分からなかったため論文を投稿しても相手にしてもらえず、論文を出すのに非常に苦労しました。Slpファミリーの機能解明のブレイクスルーとなったのが、SHD領域に低分子量G蛋白質Rab27A/Bが特異的に結合するという発見でした(J .Biol. Chem. (2002) 277,
9212-9218)。SHD領域の機能解析を行うために、まず既存の蛋白質モチーフと相同性がないか検討したところ、ラブフィリン(rabphilin)という分子のRab3A結合領域と弱いながら相同性を示すことが明らかとなりました。低分子量G蛋白質Rabは真核生物に普遍的に保存された膜輸送制御因子で、マウスやヒトにおいては60種類以上のアイソフォームが存在すること、それらは様々な膜輸送のステップを制御すると考えられています。私達は、SHD領域にはRab3A以外の何らかのRabアイソフォームが結合するに違いないと考え、マウスに存在する60種類の全てのRabをクローニングし、結合特異性を検討してみました(J .Biol. Chem.
(2002) 277, 40118-40124; J . Biol.
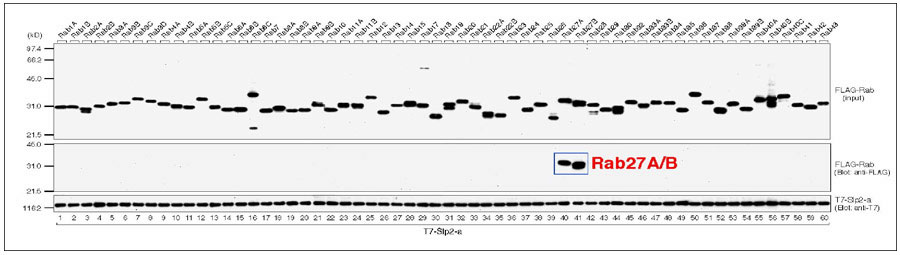
Chem. (2003) 278, 15373-15380)。その結果、驚くべきことにSlp2-aとSlac2-aのSHD領域には60種類のRabのうちRab27A及び近縁のアイソフォームであるRab27Bにのみ非常に特異的に結合することが明らかになりました(図6)。この研究成果は、Rabとの結合特異性を網羅的に解析した初めての論文として世界的に注目されました。
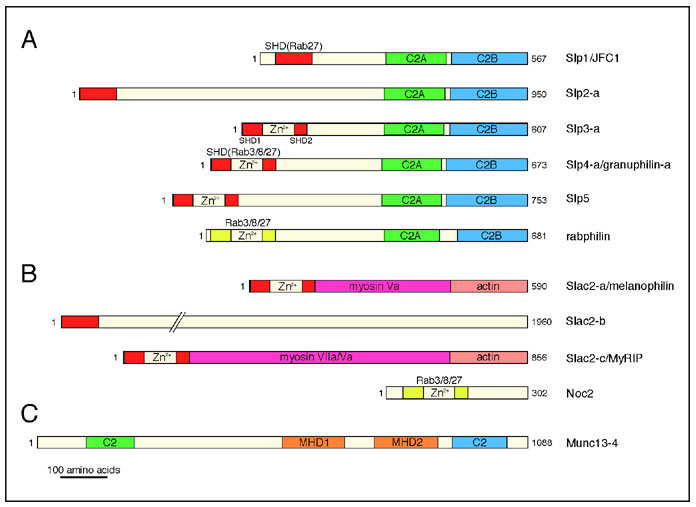
図5 Rab27エフェクター分子群のドメイン構造 (クリック)
マウス及びヒトにおいては11種類の異なるRab27Aエフェクター分子がこれまでに報
告されています。これらの分子はその分子構造からSlpファミリー(Rab27A結合ドメイ
ンSHD領域とタンデムC2ドメインを有する)、Slac2ファミリー(SHD領域をN末端側に
有するが、C2ドメインを持たない)、Munc13-4の三つのグループに分類さています。この
うちSlac2-a/cはC末端側にミオシン結合ドメイン及びアクチン結合ドメインを有します。
図6 Slp2-aと60種類のRabとの結合特異性 (クリック)
60種類のFLAG-Rabのうち、Rab27A/Bのみが特異的にT7-Slp2-aと結合する。
低分子量G蛋白質Rabは、GTPを結合した活性化型とGDPを結合した不活性化型の二つのフォームをとり、活性化型のGTP-Rabはエフェクター(effector)と呼ばれる特異的なパートナーと結合することにより膜輸送を促進すると考えられています(図7)。すなわち、Slp及びSlac2のSHD領域はRab27A/Bのエフェクタードメインとして機能するものと考えられた訳です。その後の研究により、現在ではSHD領域はRab27のエフェクタードメインとして広く認知されるようになりました。
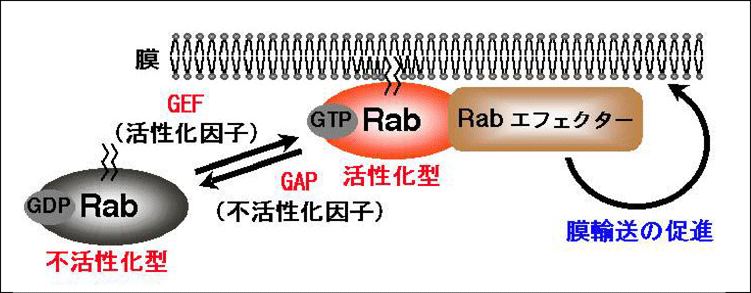
図7 RabとRabエフェクターによる膜輸送制御機構 (クリック)
一般的にRabはGDPを結合した不活性化型とGTPを結合した活性化型の二つのフォー
ムを取ります。GDP-RabはGEF/GEPと呼ばれる酵素の作用により、GTP-Rabとなり、活
性化されたGTP-Rabはエフェクターと呼ばれる結合パートナーと特異的に結合すること
により、膜輸送を促進します。また、GTP-RabはGAPと呼ばれる酵素の作用により不活
性化されます。
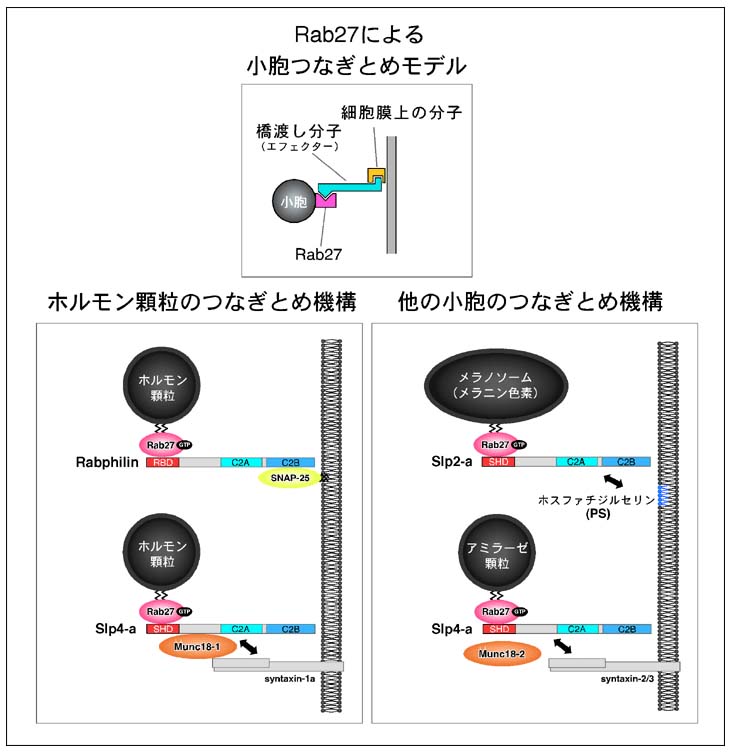
数多く存在するRabの中で、Rab27Aはヒトの遺伝病の原因遺伝子として同定された初めてのRabとしてよく知られています。2000年末にフランスのグループにより、Rab27Aの欠損によりヒトの遺伝病タイプII型Griscelli症候群が発症することが報告されました。Griscelli症候群は非常に稀な遺伝病で、この遺伝病の患者は色素異常(メラノサイトにおけるメラノソームの輸送異常)や重度の免疫不全(細胞障害性T細胞からの溶菌性顆粒の放出異常)の症状を示します。しかしながら、この当時は何故Rab27Aの欠損によりこのような多様な症状を示す(メラノソーム輸送と顆粒放出はかなり異なる膜輸送と考えられます)のかといった分子メカニズムは全く解明されていませんでした。世界中で、Rab27A以降の下流のシグナルの探索(すなわちRab27Aエフェクターの同定)が行われていましたが、私達の研究室ではシナプトタグミンの研究という全く別の切り口からRab27Aエフェクターの同定に世界に先駆けて成功しました。現在では、Rab27Aエフェクターとして11種類の異なる分子が報告されており(図5)、Rab27Aは細胞特異的あるいは組織特異的にRab27Aエフェクターを使い分けることにより異なるタイプの膜輸送(メラノソーム輸送と顆粒放出)を制御すると考えられています。私達は、副腎髄質クロマフィン細胞由来のPC12細胞をモデル系として、Slp4-a及びラブフィリンがホルモン顆粒の細胞膜ドッキングの過程に重要な役割を果たすことを明らかにしています(図8)(J. Biol. Chem. (2005) 280,
39253-39259; Mol. Biol. Cell (2006) 17, 2101-2112)。Slp4-aはMunc18-1及びシンタキシン1aと複合体を作ることにより、開口放出を起こさない細胞膜上のホルモン顆粒数(inert
docked vesicles)を増大させるのに対し、ラブプィリンは細胞膜上のSNAP-25と結合することにより、開口放出刺激に対してすぐに反応する細胞膜上のホルモン顆粒数(releasable
docked vesicles)を増大させると考えられています。さらに、私達の研究室ではRab27-Slp複合体が内分泌細胞のみならず、外分泌細胞からの分泌過程(唾液腺腺房細胞からのアミラーゼ分泌や胃の表層粘液細胞からの粘液分泌)においても重要な役割を果たすことを明らかにしています(J. Biol. Chem.
(2005) 280, 39175-39184; Genes Cells (2006) 11, 623-631)。
図8 Rab27-Slp複合体による細胞膜への小胞つなぎ止め機構 (クリック)
Rab27は成熟した分泌顆粒あるいはメラノソーム上に特異的に存在します。ここにま
ず、Rab27エフェクターSlpあるいはラブフィリンがSHD領域を介して結合します。Slp
及びラブフィリンは細胞膜上の特異的な分子(蛋白質あるいは脂質)と結合することに
より、Rab27が存在するオルガネラを細胞膜につなぎ止めます。
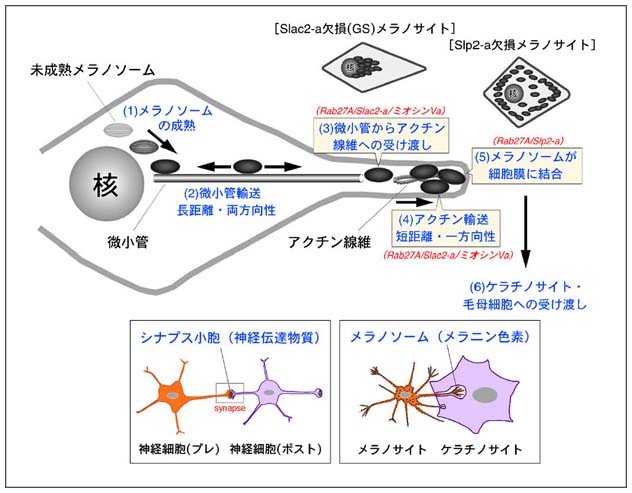
メラノサイト(色素細胞:メラニン色素を産生する細胞)と神経細胞は全く機能の異なる細胞ですが、発生学的にはメラノサイトと神経細胞は同じ神経冠に由来し、膜輸送と言う観点からみると両者にある種の類似性を見いだすことが出来ます(図9)。神経細胞においては、軸索を伸ばし、神経終末からシナプス小胞の内容物である神経伝達物質を後シナプス側の神経細胞に受け渡すことにより興奮を伝えますが、メラノサイトにおいては、メラニン色素を貯蔵するメラノソームを樹上突起を介して隣接するケラチノサイト(皮膚を作る細胞)や毛母細胞(髪の毛を作る細胞)に受け渡すことにより、肌や髪の毛の暗色化が引き起こされます。つまり、神経細胞とメラノサイトにおいて同じような膜輸送制御因子が機能していても不思議ではないと考え、メラノサイトにおけるメラノソーム輸送の分子メカニズムの研究にも着手することにしました。メラノサイトにおけるメラノソーム(メラニン色素含有小胞)の輸送は図9に示すように行われます。私達の体が太陽の光を浴びると、皮膚の基底層に存在するメラノサイトが活性化され、メラニン色素が合成され、メラノソームに貯蔵されます(ステップ1)。核周辺で成熟したメラノソームはまず、長距離・両方向性の微小管輸送により細胞の辺縁部へと輸送されます(ステップ2)。次に、アクチン線維へと乗り換え(ステップ3)、短距離・一方向性のアクチン輸送により細胞膜直下まで輸送されます(ステップ4)。細胞膜につなぎ止められたメラノソームは(ステップ5)、最終的にメラノサイトの樹状突起から隣接するケラチノサイトや毛母細胞へと受け渡され(ステップ6)、肌や毛髪の暗色化が引き起こされます。メラノソームは明視野の顕微鏡でも容易に観察することができることから、メラノソーム自身が細胞内を移動することは古くから知られていましたが、その分子メカニズムの解明はつい最近まで、ほとんど解明されていませんでした。
図9 細胞レベルで見たメラノサイトにおけるメラノソーム輸送 (クリック)
詳細は本文参照。
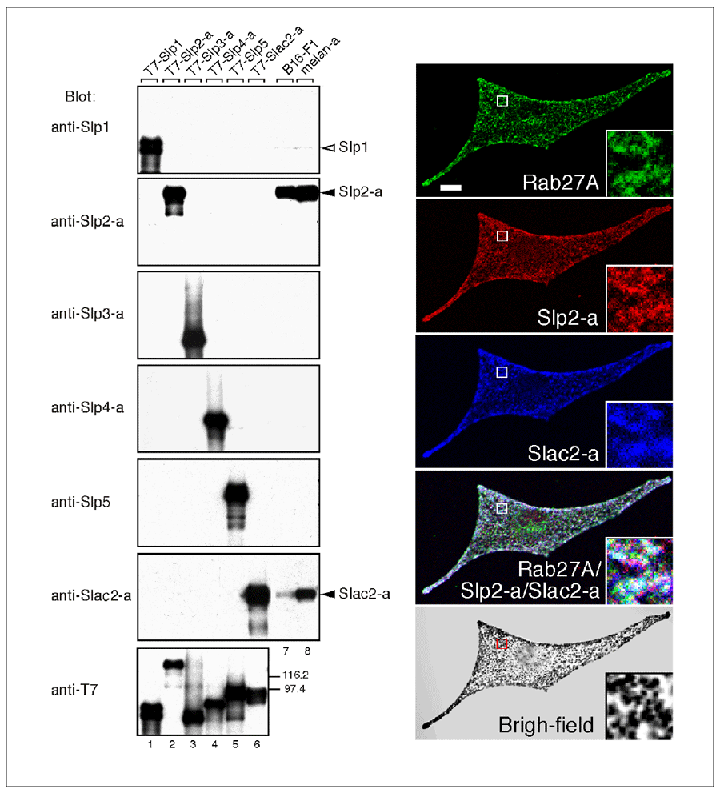
私達の研究室では、同定したRab27AエフェクターのうちSlac2-a(melanophilinとも呼ばれています)とSlp2-aの二種類がメラノソーム上に豊富に存在することを見いだし(図10)、RNA干渉法によるノックダウン実験により世界に先駆けてメラノソーム輸送の分子メカニズムの解明に成功しました(J. Biol. Chem. (2002) 277, 12432-12436; Mol.
Cell. Biol.
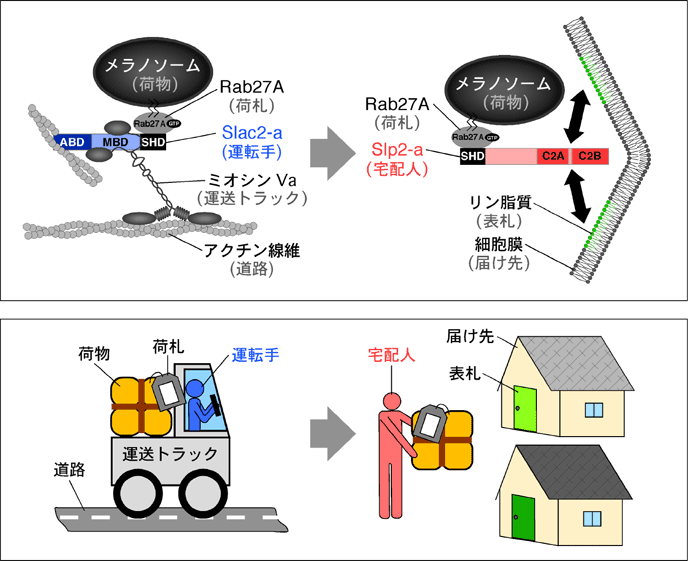
(2003) 23, 5245-5255; Nature Cell Biol. (2004) 6, 1195-1203)(図11)。Rab27A、Slac2-a、Slp2-aによるメラノソーム輸送を宅配便に例えてみると次のようになります。成熟したメラノソーム上には荷札役のRab27Aが特異的に存在し、ここに運転手役のRab27AエフェクターSlac2-aと運送トラック役のアクチン依存性モーター蛋白質ミオシンVaが結合します。この三者の複合体の形成により、メラノソームは微小管からアクチン線維へと受け渡され、その後のアクチン輸送が行われます。目的地である細胞膜が近づくと、今度はもう一つのRab27AエフェクターSlp2-aの機能によりメラノソームは届け先である細胞膜につなぎ止められます(この時、Slp2-aのC2Aドメインは細胞膜の構成成分であるフォスファチジルセリンと特異的に結合します)。本研究結果は、一つのRabが二つのRabエフェクターを連続的に用いることにより膜輸送を行うことを示した初めての事例として、当該研究分野で注目を集めました。また、Slp2-aをノックダウンしたメラノサイトでは、通常の細長い細胞形態が丸い形態に変化することから、Slp2-aにはRab27Aエフェクターとしての機能以外にも細胞極性維持の機能を持つことが明らかになっています(Nature Cell Biol. (2004) 6, 1195-1203)。このSlp2-aの機能は、メラノサイトが樹状形態を示し、隣接するケラチノサイトや毛母細胞にメラノソームを効率よく受け渡すのに一役買っているのではないかと推察されます。私達の研究室では引き続きSlp2-aによるメラノサイト細胞形態維持の分子メカニズムの解明に取り組んでいます。
図10 メラノサイトにおけるRab27エフェクター(Slp2-aとSlac2-a)のメラノソームへの局在 (クリック)
図11 Rab27A、Slac2-a、Slp2-aによるメラノソーム輸送モデル (クリック)
詳細は本文参照。
Rab27AはGEF(guanine
nucleotide exchange factor)により活性化され、GAP(GTPase-activating protein)により不活性化されると考えられていますが、Rab27Aに特異的なGEFやGAPはこれまで同定されていませんでした。私達の研究室では、Rab27A-GEFとしてAEX3/Rab3-GEPを(Mol. Biol. Cell (2006) 17, 2617-2625)、Rab-GAPドメインと考えられているTBC(Tre2/Bub2/Cdc16)ドメインを持つ蛋白質を網羅的に解析することにより二種類のRab27A-GAP(Rab27A-GAPα/EPI64及びRab27A-GAPβ/FLJ13130)を同定することに成功しています(J. Biol. Chem. (2006) 281, 31823-31831)。Rab27A-GAPαはメラノサイトにも発現しており、Rab27A-GAPαの過剰発現はメラノソーム輸送を強く阻害することから、美白化粧品の分子標的としても応用できるものと期待しています。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}